 ad clásicamente se refiere a una reacción inmune exacerbada que produce un cuadro patológico causando trastornos, incomodidad y a veces, la muerte súbita. Tiene muchos puntos en común con autoinmunidad donde los antígenos son propios. Las reacciones de hipersensibilidad requieren que el hospedero haya sido previamente inmunológicamente sensibilizado, es decir, que haya sido expuesto a lo menos una vez a los antígenos en cuestión. La clasificación en cuatro grupos distintos fue expuesto por P. H. G. Gell y Robin Coombs en 1963.[1] En la década de 1930 Coombs sistematizó estas reacciones de acuerdo al tiempo de demoraba la aparición de los síntomas y la dosis de desafío. Esta clasificación no solamente apuntaba a la cinética de las reacciones, sino que a los mecanismos involucrados y ha sido fundamental para orientar la terapia y conocer los mecanismos.
ad clásicamente se refiere a una reacción inmune exacerbada que produce un cuadro patológico causando trastornos, incomodidad y a veces, la muerte súbita. Tiene muchos puntos en común con autoinmunidad donde los antígenos son propios. Las reacciones de hipersensibilidad requieren que el hospedero haya sido previamente inmunológicamente sensibilizado, es decir, que haya sido expuesto a lo menos una vez a los antígenos en cuestión. La clasificación en cuatro grupos distintos fue expuesto por P. H. G. Gell y Robin Coombs en 1963.[1] En la década de 1930 Coombs sistematizó estas reacciones de acuerdo al tiempo de demoraba la aparición de los síntomas y la dosis de desafío. Esta clasificación no solamente apuntaba a la cinética de las reacciones, sino que a los mecanismos involucrados y ha sido fundamental para orientar la terapia y conocer los mecanismos.CLASIFICACION DE COOMBS Y GELL

TIPO 1 - inmediata (o atópica, o anafiláctica)
Artículo principal: Alergia
La hipersensibilidad tipo 1 es una reacción alérgica provocada por re-exposición a un tipo específico de antígeno referido como un alérgeno[5] La exposición puede haber sido por ingestión, inyección o por contacto directo. La diferencia entre una respuesta inmune normal y una hipersensibilidad de tipo 1 es que las células plasmáticas secretan IgE. Esta clase de anticuerpos se unen a los receptores para la porción constante (Fc) del anticuerpo sobre la superficie de los mastocitos tisulares y basófilos circulantes. Al cubrirse estas células con IgE son sensitizados al momento de la aparición inicial del alergeno. Con subsecuentes exposiciones al mismo alergeno, hace que las IgE se entrecrucen en la superficie celular de células sensitizadas, resultando en una desgranulación y secreción de mediadores farmacológicamente activos, tales como la histamina, leucotrieno y prostaglandina. Los principales efectos de estos productos son la vasodilatación y la contracción del músculo liso.
Este tipo de reacción puede ser localizada o sistémica. Los síntomas varían de una irritación leve a la muerte súbita por anafilaxia. El tratamiento generalmente involucra el uso de epinefrina, antihistamínicos y corticosteroides.
Dermatitis atópica (eccema)
Urticaria (habones)
Eosinofilia
Penicilina
TIPO 2- dependiente de anticuerpos
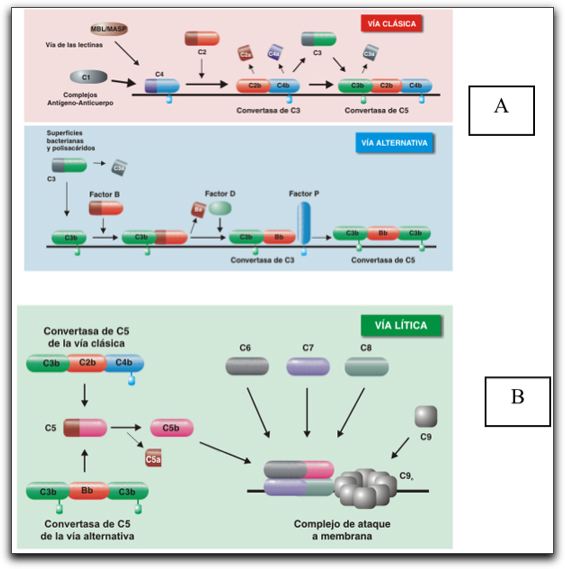
En la hipersensibilidad tipo 2, los anticuerpos producidos por el sistema inmune se unen a antígenos en la superficie misma de las células del paciente. Los antígenos así reconocidos pueden ser de naturaleza intrínseca (son parte innata de la célula del paciente) o extrínseca (absorbidas a la célula durante la exposición a un antígeno extraño, posiblemente una infección por algún patógeno. Estas células son reconocidas por macrófagos o células dendríticas que actúan como células presentadoras de antígeno, lo que causa que las células B respondan produciendo anticuerpos en contra del susodicho antígeno. Un ejemplo es la reacción a la penicilina, en el que la droga se une a los eritrocitos causando que éstas sean reconocidas como extrañas para el cuerpo. Ello hará proliferar las céluas B junto con la secreción de anticuerpos en contra del medicamento. Los anticuerpos de tipo IgG e IgM se unen a éstos antígenos formando complejos que activan la vía clásica del complemento iniciando una secuencia que terminará con la eliminación de las céluas que presentan los antígenos extraños, causando lisis y muerte celular. Ese es el proceso regular de eliminación de patógenos, volviéndose peligroso para el hospedador si el proceso se activa en conrta de sus propias células. La reacción puede durar horas o días en completarse.
 TIPO 3- Complejo inmune
TIPO 3- Complejo inmuneEn la hipersensibilidad tipo 3, se forman en la sangre complejos inmunes solubles, es decir, agregados de anticuerpos IgG e IgM), que son depositados en varios tejidos (típicamente la piel, riñón y las articulaciones donde disparan una respuesta inmune fundamentado en la vía clásica de la activación del complemento. Hay dos etapas relacionadas al desarrollo de complejos inmunes, primero el complejo se forma cuando los anticuerpos IgG e IgM se unen al antígeno, luego de lo cual, los complejos se tornan de mayor tamaño los que pueden ser eliminados del cuerpo. Es en la primera etapa de esta formación que no es posible eliminar estos complejos antígeno:anticuerpo del organismo, por lo que son esparcidos y depositados en los tejidos mencionados. La reacción toma varias horas hasta días para desarrollarse.

TIPO 4 - Mediada por células (Hipersensitividad Tipo Retrasada o Tardía, DTH)
Artículo principal: Inmunidad celular
La hipersensibilidad tipo 4 es frecuentemente llamada tardía, pues a la reacción le toma 2 o 3 días para instalarse. A diferencia de los otros tipos, no es mediada por anticuerpos, sino por células inmunes.
Los linfocitos T CD8 y CD4 cooperadores reconocen los antígenos en un complejo con el complejo mayor de histocompatibilidad tipo I y II. Las células presentadoras de antígeno en este caso son los macrófagos que secretan IL-12, el cual estimula la proliferación de más linfocitos T. Los CD4+ secretan también IL-2 e interferón gamma, estimulando aún más la liberación de citocinas, de ese modo mediando la respuesta inmune. Las células CD8 destruyen las células diana al entrar en contacto con ellas mientras que los macrófagos activados producen enzimas hidrolíticas y, ante ciertos patógenos intracelulares, se transforman en células gigantes multinucleadas.
BIBLIOGRAFIA
http://images.google.com.mx/imgres?imgurl=http://www.ebrisa.com/portalc/media/media-S/images/00016194.jpg&imgrefurl=http://www.ebrisa.com/portalc/ShowArticle.do%3Bjsessionid%3DD2C72F93DED48F7219BCA41EB2421C64%3FarticleId%3D93882&usg=__dk7_jqGGsBGPdkHGUMjSHh2WWX0=&h=878&w=654&sz=54&hl=es&start=11&um=1&tbnid=Sa9j-H3UaUY_xM:&tbnh=146&tbnw=109&prev=/images%3Fq%3DHIPERSENSIBILIDAD%26hl%3Des%26sa%3DN%26um%3D1
http://es.wikipedia.org/wiki/Hipersensibilidad


{kind=link}
{kind=link}
{kind=link}
{kind=link}